2015-11-01 14:03:46
按: 本文有些观点值得商榷, 但读了之后颇能引人思考, 不失为一篇好的科普文. 我忽然想到现今的有些生物问题是不是仍然可以从本文所说的方面考虑一下呢? 比如癌症. 研究得越多真相仿佛越模糊, 各种致癌物质五花八门, 加工肉被世卫组织列为最高风险致癌物. 这是不是说明我们犯了与以前同样的错误呢?
前言
在以前本小组的讨论中,我曾提到“比如你如果不理解所有生物共有的膜渗透能量供应化学原理,你就根本无从评价任何生命起源的理论。”
在一篇题为“为什么还有这么多人相信进化论?”的帖子中,蒙hongdou网友发信询问,所以这篇文章既是一篇泛泛的科普,也是对这个问题的具体介绍。
本文并非全部原创,其中有大量数据、理论和史料来自于英国生物化学家、科普作家Nick Lane的著作 Oxygen 和 Mitochondria。这两本书都是非常好的科普读物,可惜没有中译本。大致引用了他1/3,自己发挥了1/3,以及扯远了1/3。特此声明和致谢。
- 曼斯.雷德
一 直觉与反直觉
直觉(intuitive)和反直觉(counterintuitive)是科学讨论在描述一个科学理论或者发现的时候,经常使用的二分法。这个叫法本身并不那么科学和严谨,但是其中的意味却是无限深长的。
既然不严谨,我也不去定义它,只看范例:
最简单清楚的直觉理论,在古希腊科学有很多范例。比如“刚性物体排开的水量等于它的体积”。比如欧几里得平面几何中的公设和简单定理。“凡直角都相等”,“两点之间直线最短”,初学几何的人,都会想这不是废话吗。等到用这些废话为基础武器,逐步分解,证明了其他复杂得可怕的理论,我们才知道废话的精辟之处。这就是直觉理论最原始的特征:不言而喻。然而,即使在这种萌芽时代的智慧中,也埋下了“反直觉”理论的种子,那就是仅仅“篡改”了一条欧几里得公设,就闯出一片新天地的罗氏几何。

牛顿第三定律是另一个直觉理论的例子:作用力必然导致反作用力。这看起来也是不言而喻的废话。但牛顿第一定律就不是了:“物体将保持它的运动状态,直到外力改变它”。这是一个相对原始的反直觉理论。在今天,初中物理课修完的同学也许会不同意这是反直觉的。但是在牛顿的年代,这却是彻头彻尾的反直觉。无数牛顿之前的大学者,都理所当然地认为物体要保持运动,必须不断注入外力。这才是费劲力气保持各种物体运动的古代人类的“直觉”。另一个直觉对阵反直觉的著名例子是伽利略对阵亚里斯多德(我不叫它“比萨斜塔实验”,是因为这个传说本身是有误的。但是双方的论点是明确无误的)。“更重的物体下落更快”,这样既简单又错误的直觉,连亚里斯多德这样的智者都不能免俗。可以保证,即使在21世纪的今天,一个纯文盲多半仍然会有这样的直觉。
直觉与反直觉是相对的,也是可以培养的。经过一定科学训练的人(比如说读完初中物理),他的直觉会超越两足动物的局限,随着训练的程度达到新的精确度,比如说把牛一定律变成他的直觉。但一个人具体的生活年代和环境,并不反映他应有的直觉水平,或者他的直觉正确程度。比如我95岁的奶奶(文盲),直到今天仍然不同意我说的地球围着太阳转。比如美国仍然有20%以上的人认为地球生命历史不超过4000年。
十八、十九世纪是直觉型科学理论达到顶峰的年代。能量守恒定律(在不久以后就将被进阶的“反直觉”理论给河蟹了),热力学第二定律(“一个房间不花力气整理只能越来越乱”),在它们刚刚被提出的年代,文盲仍然会认为是反直觉的,受过良好科学训练但非专业的头脑,一听到就知道是正确的。而真正的前沿研究者则是醍醐灌顶:“这么简单明白,怎么我就没有归纳出来呢?”非常精彩,非常辉煌,但在物理学上,一般人类直觉能够企及的高度也就到此为止了。接下来的爱因斯坦、海森堡、薛定谔,一个比一个猖狂地反直觉。相对论刚刚提出的时候,不要说普通人,就是顶尖物理学家,也认为爱因斯坦简直是在说胡话。对今天世界上99%的人来说,相对论和量子物理仍然是胡话。然而我们现在都知道,他们是正确的。因为广岛真的被炸掉了,我们车上装的GPS也真的用上了相对论校正。
“反直觉”的优势在现代物理学中很清楚。能够以普通人类直觉发现的真理,基本都被发现了。剩下的真理必然都很古怪难以接受。在这样的高度还去依靠直觉,多半都要犯错误。比如爱因斯坦的“上帝不掷骰子”,听起来很美好很庄严,结果证明错误。同样是老爱的坚信宇宙密度常数严格等于1,宇宙会无限震荡,听起来很和谐很完满,从现在的证据看来,也是错得离谱。即使是无数物理学家(再一次包括老爱)追求的圣杯:统一场理论,也是一个高级的直觉型假说,前景仍然不妙。总之在现代物理学中,寻求反直觉的答案,克服直觉障碍去接受和欣赏反直觉的理论,反而被养成了研究者的本能。看看霍金有多少胡话(姑且不说他的胡话站得住脚不),以及被学界和起哄界怎样异口同声地热捧。
然而在生物学中,直觉与反直觉的力量对比就完全是另一回事了。
达尔文的进化论,是生物学中开天辟地的反直觉理论。在著名的钟表匠辩论中,神创论者洋洋得意的雄辩(Paley主教:“我们看见地上有一个精致的怀表,自然能肯定某处一定有一个钟表匠!”),首先是一个蹩脚的比喻,然后不过是一个粗浅短视的人类直觉。如果放到现代物理学那种气氛中来看,连认真驳斥的价值都没有。然而这种直觉式的大脑当机,却能跟进化论翻翻滚滚地斗上一百多年。直到今天,仍然妨碍大众在生物学上入门,仍然堂而皇之披上科学的外衣来反科学,仍然有少数严肃的学者在掉进相同的直觉陷阱(Fred Hoyle,天文学家和数学家:“生命起源于地球的概率就像大风吹过机械垃圾场组装出一架波音747的概率”。这句话被各种神棍肆无忌惮地篡改,主语直接变成了“进化论”。实际上Hoyle并不反对进化论原则,只是在概率计算上有异议,从而认为地球生命必然有地外来源,才有时间来满足他的概率计算结果。他的领域是生命起源)。这篇文章的主题不是跟他们叫板,所以这里先暂且不提,后文再加讨论。
并不是说,科学家就不会被直觉蒙蔽。前面曾提到,反直觉天才如爱因斯坦,也曾连挖三个直觉的大坑。只是在生物学中,直觉理论的顽固性来得特别的强,而对反直觉理论的接受来得特别的艰难,不管大众和专业领域都是如此。进化论就曾有直觉与反直觉的争论,即是用进废退学说和自然选择学说。在那个对遗传的物质细节一无所知的年代,用进废退理论的描述来得如此自然。“长颈鹿需要吃高处的叶子,伸着伸着脖子就变长了”,直到我上小学《自然》的时候,我的老师仍然是这么讲的!近两百年之后都还是这样,可想而知达尔文主义在初生年代的四面楚歌。话说回来,即使艰深繁杂如生物学,仍然是科学。科学的伟大就在于在科学的竞技场中,我们可以肯定真理必胜,即使前面被拍倒一百次,走上一千个曲折。人生中其他的事情,不管是政治、感情、事业还是道德正义,我们都不能打这样的包票。这也是科学对我的终极吸引力。
这篇文章要介绍的化学渗透理论,就被誉为二十世纪最伟大的“反直觉”生物学发现,虽然它并不为公众熟知。这里所说的直觉,并非一般常识,已经是二十世纪生物学家和化学家中非常精深的理论直觉了。然而当直觉走入死胡同时,一个“怪异”程度不下于相对论或者进化论的新理论蹦了出来。它在初期所受到的不公正待遇,以及它最后的辉煌成功,都是可以相提并论的。
二 背景
作为一篇关于生物化学的科普文章,我决定不写公式,不写化学反应方程式,不画图表。希望严谨的读者能原谅这点小小的文字技巧上的虚荣。
二十世纪最伟大的生物学发现(甚至可以夸大到“科学发现”),公认是沃森和克里克确定生物遗传物质是DNA,以及DNA遗传的生物化学机理。这是一个标准的直觉式发现。沃森在搞清楚DNA的双螺旋结构之后不到两个星期的时间内,就洋洋自得地告诉克里克:“我想我发现了生命的秘密。”这种从结构到功能突破性的跳跃,来自于沃森作为化学家的直觉:分子结构宣示分子功能。双螺旋意味着互为模板以及无限扩充。四个碱基两两配对意味着精确、无损的信息复制。从碱基翻译到氨基酸的简单语法意味着有限、简练的基本指令集。生物遗传的稳定性和精确性在DNA的结构中得到完满解释,从一个生物化学家的视角来看,这个跳跃简单明晰不言自明,足以让沃森和克里克之外的所有人把自己的脑袋拍肿。这个理论的被接受程度,也是犹如星火燎原,一夜之间清洗了所有研究者的大脑,改变了整个生物学研究的版图。 这个发现还运用了一个更深层次、更广泛的科学直觉现象。那就是自然科学的层次结构。
自然科学中的各个领域,并不是齐头并进的,其中一些学科是另一些的天然基础。最主要的一个层次系统是这样的:
物理学--> 化学 --> 生物学 --> 医学,人类学,心理学 --> 社会科学
越前面的越基础,每一种科学也许人类开始认识的时候是独立的,但追究到深处,你总是会发现你需要前面一层的大量知识才能产生认识突破。这样抽象地说也许很难理解,看看例子:
化学元素周期表可以算是独立化学研究的殿堂了。但即使在周期表完成之日,化学家们也很难解释,为什么元素会表现出那样的化学性质,而且周期性地回归到相似的化学性质(比如氧族元素),而且这个周期又在不断变化。单纯在化学的领域折腾,哪个理论也解释不了全部的事实。
然而,一旦物理学中的原子结构研究成熟,周期表的问题就犹如庖丁解牛,迎刃而解了。化学性质决定于外层电子配置。元素表周期就是外层电子周期。相似的化学性质就是相似的外层电子配置。基于这种理解上的飞跃,一系列重要的化学问题,比如化学键的本质,化学反应能的研究,纷纷在原子物理的层面得到突破。这种化学问题在物理层面上的理解,我把它叫做“触底”。一个领域的研究一旦触底,透彻的理解和新的发现就指日可待。
再来看一个比较贴近生活的著名例子:
抑郁症,作为心理学中的课题,一直困扰着病患和心理学家。在二十世纪上半段,弗洛伊德主义盛行于西方,抑郁症完全用心理分析方法来解释,童年经历、未实现的欲望、父母责任,一系列五花八门的弗洛伊德式学说和疗法,对这个问题也无可奈何。但是当相关的生物学产生一次“触底”:发现锂制剂及其他药物的神经生物作用后,心理学也同时在生物学上触底了。90年代一桩有名的诉讼案揭开了弗洛伊德主义在西方崩溃的序幕。加利福尼亚一位长期受抑郁症折磨的中年妇女,接受了五年以上的聊天式弗洛伊德疗法,花了几万美元,从未有任何缓解。后来开始服用抗抑郁新药物“百忧解”,四个星期后痊愈。为此她愤而起诉心理医生,最终闹成了全美新闻。现在,百分之九十以上的临床抑郁症案例都用药物治疗,大多有不同程度的缓解。
沃森和克里克的成功,很大程度上也归功于生物学在化学上的触底。因此,在现代分子生物学研究中,一旦研究者逼近纯粹的化学解释,他们总是能闻到成功的气息迫在眉睫。这也就成为了现代生物学者新的直觉。
分子生物学的研究有三个宏观面:物质、信息、能量。物质是共有的基础,而信息和能量是两个不同的研究方向。DNA遗传理论的辉煌成果,把公众的视野完全集中在信息的方向,甚至学界本身也产生了认识上的偏斜。在很多人的心目中,分子生物学就是信息生物学。生物的基因组就是编码库。生命活动就是这些信息互动的总汇。而另一个宏观面:细胞能量学(或者能量生物学),在公众的认识中就非常陌生了。其实二十世纪这个领域中的认识和发现,精妙深刻和激动人心之处并不逊色于信息面。然而其中的坎坷和争执,与信息生物学中的盛世场景就大异其趣了。
二十世纪能量生物学的中心问题就是细胞能量代谢的化学本质。或者说有氧呼吸作用的化学本质和细节。直觉的方向感,引导着生物学家们从两个方向出发,向中央逼近这个课题,来试图完成拼图。就像从两端开始拼成一座大桥。截止到五十年代,大桥好像已经逼近了接龙点。已经取得了如下的成果:
- 有氧呼吸的本质和有氧燃烧并无不同,都是氧化-还原反应。有氧呼吸的氧化剂是氧气(O2),还原剂可以简化认为是葡萄糖(C6H12O6)。
- 不像自然失控的普通有氧燃烧,有氧呼吸在细胞中是一步步精确控制的。而且涉及一系列复杂的生物催化剂。
- 有氧呼吸的场所是细胞中的线粒体。线粒体内膜上镶嵌着一系列的微型催化工厂:细胞色素酶蛋白质综合体。在这些不同的催化工厂,一个个电子被从糖类上剥离,中间产物分子逐步传递,同时释放化学势能,而氧得到这些电子并结合糖中的氢原子,逐步生成水。
- 所有生物细胞中通用的能量货币是三磷酸腺苷(ATP)。它是一种高能形式的分子。对应的低能形式是ADP。
- 所有耗能的生命活动都在消耗ATP,把它们转变为ADP(以及失去一个磷酸基团:Pi),并利用转变放出的能量。因此,有氧呼吸的作用就是这些活动的反过程:制造能量,用来把低能的ADP转变为高能的ATP。
- 制造ATP的场所在ATPase,另一种镶嵌在线粒体内膜上的蛋白质综合体。这些微型催化工厂消耗ADP,释放出ATP。而且,在某些情况下,这些工厂也能反向工作,即消耗ATP制造ADP。
请注意,以上的成果我分成了两组。因为中间还缺乏一个步骤,细胞色素酶和ATPase是不同的蛋白质综合体,它们在线粒体内膜上的分布,物理上是互相隔绝的。而所谓化学反应,在分子层面上是100%需要物理接触的。那么,能量是如何从细胞色素酶传送到ATPase上的呢?这就是所谓接龙点。
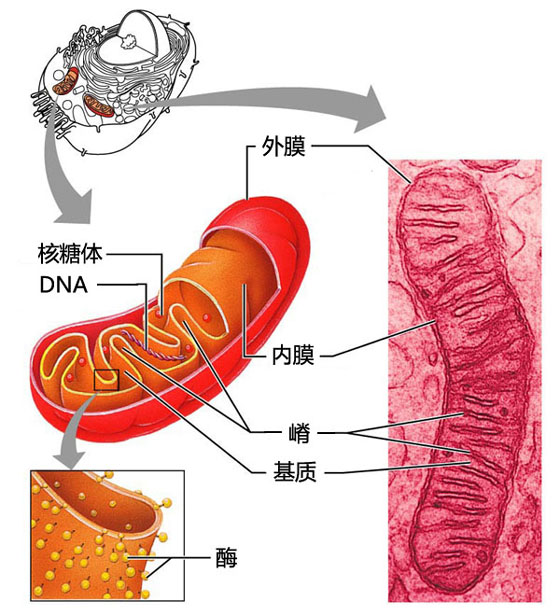
前一组是分子生物化学,后一组也是分子生物化学,中间还缺乏一个链条。你的直觉是什么?当然还是分子生物化学。我们需要找到一种媒介分子,从细胞色素酶工厂的氧化还原反应中产生,在ATPase工厂中消耗,携带一个高能状态的化学键。暂且把它叫做媒介分子X。
四五十年代的能量生物学界弥漫着乐观情绪,大厦只差一步就能建成,科学家和研究小组之间展开了寻找X分子的竞赛。然而,足足找了20年,提出的候选分子不下十几种,一个个都被研究和实验否定了。在寻找的过程中,随着对整个过程细节研究越来越深入,一系列看起来很琐碎,但是莫名其妙让人不安的问题冒了出来:
- 我们已经知道了ATP和ADP是能量通用货币的正负两种形式。在化学上,一个不言自明的直觉理解是高能状态不稳定,低能状态稳定,所谓“水往低处流”。如果把纯粹的ATP放在生物体外的溶液中,它们就会很自然地迅速转化成ADP,并释放热能。然而在生物细胞中,ATP和ADP的比例通常都是非常高能化的,ATP占了90%以上。这种全局的高能状态,需要持续的能量供应来维持。细胞中有氧呼吸的强度(用糖和氧的消耗速度来测量)变化很大,有时候相当衰弱。然而不管有氧呼吸的强度怎样,ATP的比例都维持在稳定的高位。这就像一块巨石悬在墙头,不管你用不用力撑着,它偏不掉下来。在化学的视野中,这是不可理喻的场景,然而又找不到是什么东西在支撑着它。
- ATP的产出和葡萄糖的消耗,这个反应方程式,在实际测量中居然配不平。学过中学化学的,都应该知道什么叫配平反应式。几分子的A和几分子的B反应,生成几分子的C。一旦配平,比例就是固定的。这是“数学”,所有科学的基础真理。然而测量表明,一分子的葡萄糖被完全氧化,生产28-38个ATP分子,之间任何数字都有可能,但大多数时候接近下限。在化学反应方程式中,什么叫28-38?配平需要的是整数!这又是不可理喻的反化学场景。测量实验反反复复地做,结果一直都是那样。
- 化学反应的能量和物质是守恒的,也就是说,反应式链条确立之后,前面的反应发生,后面的反应就该受驱动同时发生。前面氧化消耗葡萄糖,后面就该驱动生成ATP。这在化学术语中叫做coupling,或者耦合。消耗葡萄糖的过程和生成ATP的过程是耦合的(先忽略前面提到的配不平问题)。然而一些特殊的化学物质,可能扰乱耦合的稳定,这叫做“解耦”(uncoupling)。有氧呼吸的解耦是普遍存在的,在有些物质参与时,氧化葡萄糖的过程完全正常进行,ATP生产完全停顿。就像皮带轮系统断掉了皮带,前面的轮子空转,后面的不理睬。在化学上,这是可以接受的。因为解耦物质总是有一种化学性质,能够扰乱连续反应链中的一个环节(比如说,如果有X媒介分子,某种解耦物质倾向于和X分子结合使其失去浓度,破坏了能量传递,当然就能解耦。)但是现在的问题在于,实验证明能解耦有氧呼吸的物质,如水杨酸(阿司匹林),白喉霉素,摇头丸,化学组成和化学性质五花八门,简直找不到一个共同点。在寻找X分子失败的过程中,生物学家们对解耦物质寄予厚望,因为一旦发现了解耦物质的共同化学性质,多半就能推导出X分子到底是什么–因为这是大桥上唯一不为人知的部分。然而现实是,解耦物质发现得越多,就越找不到化学共同点。
天才的美国物理学家、科普作家费因曼,曾经对物理学中的某些困境有一个令人发笑的描述:“如果你还没有彻底被搞昏,只能证明你不懂这个领域。”在五十年代末,对有氧呼吸的研究和X分子的身份之谜,就完全是这样的场景。越是研究得透彻,理论和现实就越是自相矛盾,不可理喻。每一种假说都有不可治愈的痛脚,每位前沿研究者都不知道到底是哪里出了问题。乐观已经被混乱取代。
三 彼得·米切尔:泵,水坝和泄洪道

化学渗透的发现者彼得·米切尔从里到外都散发着离经叛道的天才气息。这个家伙家财万贯(他的叔父拥有一个建筑业集团公司),在英国二战后的困顿年月开着劳斯莱斯招摇过市。后来当他因为化学渗透的论战几乎被踢出学界时,干脆自己出钱建了一个研究所继续战斗。他在二战中由于身体原因没有服役,而当时英国能量生物学的权威克林(细胞色素酶的发现者)私下对别人说:“英国最好的科学头脑躲过了子弹”。要知道,当时米切尔才二十多岁,研究方向不是细胞能量学,而只是涉及细菌的生物学,跟克林的方向(也就是研究有氧呼吸原理)差得很远。而且他读博士读了七年才拿到学位,因为他的论文审查人多次认为他的论文都是胡说八道。同样是克林向别人解释:“米切尔的独创性对这些学者来说太多了。”
米切尔伶牙俐齿,态度粗暴,披着一头效仿青年贝多芬的长发,弹得一手好钢琴,而且和贝多芬一样很早就聋掉一只耳朵,让他跟人争吵的音量变得特别的大。一般古板的学者很难喜欢这样一个人。克林在早期的慧眼只能解释为天才识天才。
在能量生物学界因为种种化学上的不可调和而苦恼时,米切尔却从一个意想不到的方向:经典细胞生理学,接近了这个问题。
他早期的研究课题是细菌,特别是细菌的主动运输,即穿过细菌的细胞膜,营养物质和废物在受控状态下进出。我们知道在孤立的情况下,物质的渗透总是从高浓度向低浓度的方向进行,而细菌的细胞膜是脂质的半透膜,水溶性物质被阻挡,但膜上有各种特化的蛋白质通道,选择特定的物质允许进出,其严格的对应性好比特定的生物酶对应特定的催化对象。半透膜的特性加上细菌消耗能量的主动运输(当时是如何消耗能量的还完全不知道),使得细菌可以逆着浓度运输物质,比如把营养物质从低浓度的外界环境运进高浓度的内部。
对于细菌主动运输的研究,使得米切尔对生物膜导致的化学物质浓度聚集有了特别的敏感性。把细菌的主动运输过程倒过来看,如果逆浓度运输耗能,那么逆浓度运输造成的浓度聚集本身就是一种势能,好比你把石头推上坡花了力气,坡上的石头也就有了势能,随时可以滚下来释放能量。这就是米切尔的突破点。
1961年米切尔在爱丁堡发表的论文,充满自己生造的词汇,类似玄学的逻辑讨论和无证据的猜测。首先,他运用福尔摩斯式的逻辑:“当排除了其他所有可能的答案,那么剩下的可能性,不管看起来有多么不可能,都是正确的答案。”。X媒介分子的寻找,化学上每一块石头都翻起来看过了,既然还没有找到,那么答案是什么?X分子不存在。那么化学上的两套反应是如何接触的呢?能量是怎么从细胞色素氧化工厂传递到ATPase呢?米切尔说,那不是分子遇上分子的化学,那是化学渗透(chemiosmosis)。这是一个他生造的词,含有两个词根:chem(化学)和osmosis(渗透),其实这个过程的实质既不化学,也不渗透。它的含义如下:
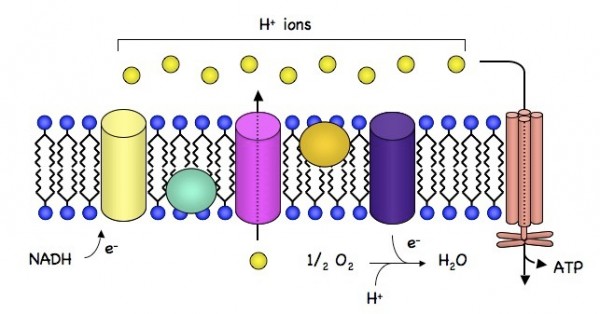
- 葡萄糖在细胞色素酶工厂中逐步氧化。这些蛋白综合体微型工厂,都镶嵌在线粒体内膜上,实质上是一些通向膜外的蛋白质孔道。它们没有合成什么高能的中介分子,作用不过是像一些泵,把细胞内的氢离子(H+,也就是质子)泵到膜外去了。这个过程需要耗费氧化葡萄糖放出的能量,因为我们下面就会看到,线粒体内膜的外部聚集着高浓度的质子。这是一个逆浓度运输。
- 由于这些质子泵的工作,大量质子被泵出去,又被限制在线粒体内膜和外膜之间的狭窄区域里,于是形成了一层薄薄的H+聚集区。H+的浓度,化学上有个很简单的量度,就是pH值,即酸性。因此内外膜之间的区域就形成了一个质子“水库”,具有酸性和正电荷。
- 线粒体内膜就像是水坝。它也是脂质膜,对于水溶性的质子来说是不通透的,因此外部质子浓度虽高,但不能自然渗透进来。记住我们前面的结论:逆浓度运输造成的浓度聚集本身就是一种势能。
- ATPase就是泄洪道。它也是镶嵌在线粒体内膜上的蛋白质综合体,也是内外相通,但受控的蛋白质孔道。它利用的不是什么神秘的X分子,而是质子的回流。质子浓度势能的释放放出能量,这些能量就被用来生产ATP。
对于那些毕生浸淫在经典生物化学,不太熟悉生物膜动力的能量生物学家来说,这简直是玄而又玄的天书,没有比这更奇怪的场景了。不去使用一个简单的中介分子完成化学链路,而是转了这么多的弯,使用这么诡异的机理。再加上米切尔生造了大量的词汇来描述他的理论(比如米切尔把“质子水库”的酸性和电势能合起来叫做proton motive-force,“质子动力”,认为这是一种所有生物的基本驱动力。听起来真像某种民科发明的词汇),整个文章写得既玄秘又武断。而且,他没有什么实验证据来支撑他的学说,基本上全是假设、推理,或者干脆忽略那些暂时解释不了的障碍。
所有这些原因,使得米切尔的论文在1961年的能量生物学界受到的待遇,差到不能再差:有些人读完第一页就扔一边。有些人认为“他终于疯了”。有些人开口就骂,认为他不尊重整个学界前人的辛苦工作。或者“不做实验,不是化学家,偏要谈最艰深的化学问题,而且他的语言不是化学。”当时的两个权威:美国的Racker和瑞典的Slater,都是很优秀的学者,一开始还不像其他人那样对待他,试图去发现米切尔到底要说什么。但Racker在讨论中完全被米切尔的外星词汇搅昏,最终放弃了讨论。而Slater和米切尔的争辩火爆升级,在一个研讨会上,Slater被米切尔尖酸刻薄的口才气得“在走廊上乱跳,白色的胡子上下飞舞”。
米切尔成了英国科学界的笑料,1963年因为“脾气大造成的胃溃疡”退出了爱丁堡大学(其实是学校当局劝退他免得丢人),回到他的乡间别墅去养病。他花了两年时间舔伤口,顺便把乡间别墅翻修成了“音乐怪人”的巢穴,找到了一个相信他,而且有很强生物化学实验能力的女助手Moyle,在1965年卷土重来,成立了自己的研究所,一定要和整个能量生物学界斗到底。
四 思想的凯旋

在1961年米切尔抛出他的重磅炸弹时,他确实没有什么实验基础,这也是大多数学者本能上不接受他最主要的原因。然而这个独辟蹊径,自己宣示一个新体系的假说,却是建立在米切尔对已知科学事实和实验结果深入分析的基础上。透过他奇特的视角和迷雾一样的语言,越来越多的人认识到,米切尔的化学渗透假说像一块奇形怪状,但是边边角角都恰好合适的拼图板,解释了有氧呼吸研究中众多的不解之谜:
- 首先就是膜。原核细胞生物(细菌和古菌)都有脂质细胞膜。而真核细胞生物中,线粒体有氧呼吸制造ATP,叶绿体的光合作用也制造ATP,这些跟能量相关的细胞器也都有脂质膜。在以往的研究中,生物膜被当做是仅仅结构上的东西,没有人从功能上意识到它的重要性,化学渗透学说解释了为什么ATP的制造场所无一例外都有生物膜。因为化学渗透的全部动力就来自于这个“水坝”制造的浓度隔离。
- 氧化场所和ATPase物理隔绝的问题也迎刃而解。因为化学渗透的能量传递过程就像你在一处用抽水机蓄水,而在另一处用泄洪道发电,整个水库(整个线粒体内外膜之间)环绕着所有的设备。这时候你的设备在哪里当然无所谓了。
- ATP异常的高比例问题也是这样轻易解决。只要线粒体内膜外积聚着足够浓度的质子库,ATPase就能持续把ADP转变成ATP,保持ATP在细胞内的高浓度。而这时候氧化强度怎样是无关痛痒的。水库蓄够了水,哪怕抽水机停上一阵,发电也在持续进行。
- 葡萄糖氧化量和ATP制造量配不平的问题,在这个场景下也不成其为问题。因为这样看来,制造ATP的数量根本不是受实时的氧化量控制的(实际上是受细胞能量需要控制,由其他来自外部的化学信号调节的)。就像发多少电是受电网需求控制的,跟实时的抽水量没有严格对应关系。只有长期、大体上的宏观比例,这就是28-38这个含糊数字的含义。
- 最精彩的是这个假说一下子就解决了众多解耦物质的共性。答案对化学家们来说真是郁闷,因为实在是太简单了。以往寻找共性的时候,化学家们总是在找它们的反应特性,完全没想到这些东西都有两个最简单,在高分子的生物化学中根本“不值得”考虑的属性:它们都是脂溶性的弱酸。弱酸的特点是两性化:在酸性较大的环境中表现为“碱”,跟氢离子(也就是质子)结合。在中性或碱性环境中恢复酸的本性,释放氢离子。脂溶性使得它们能够自由穿越脂质生物膜,在线粒体外表的酸性水库中拽住质子,穿越回来回到内部又释放质子。而且它们受到内外酸碱度差和电位差的往复压力,会不停地做这种来回运动。这些解耦物质就像偷渡船,把细胞色素工厂辛辛苦苦泵出去的质子又都给运了回来,只要它们达到一定浓度,外部的“水库”就被放得失去了足够势能,ATP制造当然停滞了。一个千疮百孔的大坝是蓄不了水,发不了电的。
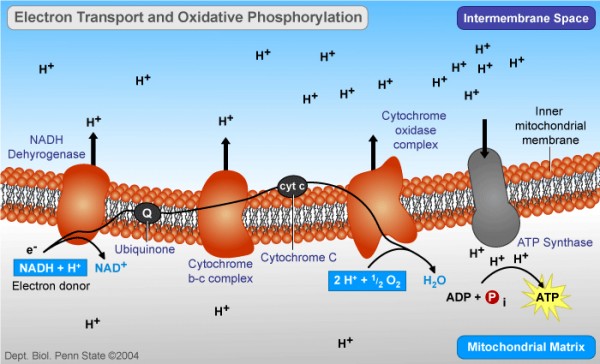
一个健康有力的科学假说,不但需要解释已经存在的疑问,而且通常都能做出一些具体、可验证的预言,描述尚未发现的现象,去引导实验科学家加以证实。1965年以后,米切尔本人和他的助手Moyle当然在做实验,其他一些脑子逐渐被米切尔持续的喋喋不休掰过来的科学家也加入了实验行列,其中包括一些米切尔以前论战得最苦涩的论敌,比如Racker。
60年代末,米切尔和Moyle首先实验证实了他自己的预言:线粒体内外膜之间不但有明确的酸性浓度梯度,和膜内还有150毫伏的电位差。不要小看这150毫伏,因为它的距离间隔仅仅是5纳米的线粒体内膜。等比例换算的话这个强度大概和大气闪电的强度差不多,比家用电高1000倍,堪称自然界强度最高的电势能场。
Racker跟着做了一个精巧的实验,他把细胞色素氧蛋白综合体从线粒体中提取出来,放进人造的脂质膜泡中,测得膜外仍然能积蓄起质子浓度。这个实验的含义是排除了有氧呼吸的消耗段:ATPase,以及ATPase自己和其他所有细胞环境循环影响质子浓度的可能性,单纯检测供应段。结果是完全符合化学渗透假说的前半段。在此之前,没人相信已经研究得十分透彻的葡萄糖氧化过程真的是“质子泵”。Racker的皈依影响了学界大量的人,风向开始转变了。
在这个阶段米切尔的促狭鬼性格开始作怪:他公开建立了一个表格,表格上记录他以前每一个批评者“皈依”的日期,Racker大人荣登榜首。其他还没归顺的学者一个个先列出来,日期留白。
下一个实验突破是瑞典的雅根多夫,他不是能量生物学家,而是植物学家。在一个科学会议上,他偶尔听到米切尔跟人论战,当时的感受是米切尔态度恶劣而且说的都是胡话—然而是他自己可以证伪的胡话,因为米切尔不但认为线粒体符合他的假说,还说叶绿体中光合作用制造ATP的机理也是同样的。对米切尔强烈的反感促使雅根多夫和他的美国同事设计了一个实验:把叶绿体膜分离出来,排除其他一切细胞环境和叶绿体内部结构,在简单的人造环境溶液中加以测试。首先用pH=8的弱碱性平衡内外酸碱度,然后加入酸液把外部pH调整到4。这相当于把米切尔的假说还原到最简结构。结果是,叶绿体膜内马上产生了大量ATP。当时雅根多夫简直不相信自己的眼睛。后来他发表自己的实验数据后,主动申请加入“归顺榜”。
越来越多的实验结果,一项项填补的细节空白,逐渐把化学渗透假说从丑小鸭变成了羽翼丰满的大怪物。进入70年代之后,能量生物学界已经全面倒戈,化学渗透成了整个有氧呼吸研究的新范式,建立在它基础上的新研究、新突破方向也爆炸性地出现了。比如美国女生物学家Lynn Margulis在1971年发表的轰动论文,证明了所有生物的线粒体前身是远古的细菌,与真核生物细胞之间是共生关系。这个成果的基础,就是线粒体有氧呼吸和细菌的基本能量代谢完全使用相同的质子泵-生物膜驱动模式(下一节会详细介绍细菌的问题)。
米切尔本人独得了1978年的诺贝尔化学奖。1961年到1978年,17年时间就从“门外汉”的状态登上了荣誉和成就的顶峰,这在诺贝尔奖的历史上是罕见的。物理、化学和生物学这三个奖项,大多顶尖科学家在做出自己最显赫的贡献后30-40年才能获诺贝尔奖。更不要说很多人终身无望。
这样的殊荣,加上米切尔嚣张的气焰,再加上获得化学奖的“化学渗透”理论是这样一个介乎化学、物理、生物三个领域之间,出乎所有人想象之外的怪异机制,使得学界的很多人大吐酸葡萄。米切尔在领奖致辞中这样感谢他们:
“马克斯.普朗克(著名物理学家)曾经说过,一个新的科学理论不是靠使权威反对者信服来取得胜利的,而是靠这些权威反对者逐渐死掉。这是非常悲观但也非常现实的箴言。我最大的成就是证明了普朗克也有说错的时候。”
1997年的诺贝尔化学奖得主沃克,他的主要成就是利用新发展的衍射晶体学技术测定了ATPase的原子级别精细结构。而这个成果,实际上是给20年前米切尔理论的登基进行了迟来的加冕。沃克昭示的ATPase精细结构,简直是一部鬼斧神工的蛋白质机器。我们在这部分最后来欣赏一下大自然炫目的工程杰作。
ATPase是一个蛋白质复合酶,微小到在一般电子显微镜下也只是隐约可见,像蘑菇一样星星点点地点缀在线粒体内膜上。它是由很多个微小的、运动的蛋白质部件构成的,各部件大体上呈同心圆环状排列。结构上,ATPase分成两个部分,一个是蛋白质排列拼合成的孔道,这个孔道直接从膜内穿透到膜外。孔道的内端是另一个部件:一个轴样的旋转头。这个旋转头像钟表里的齿轮一样,有三个固定刻度。每个刻度之间角度相差120度。外部的高浓度质子“水库”产生的势能促使质子从孔道内滑落,大致每三个质子的滑落的能量会让旋转头转过120度,即一个刻度,10个质子穿过完成一个循环。这个蛋白质旋转头每次转动一个刻度,它的分子张力都让跟它接触的ATP、ADP分子化学键生成或者打破。第一个刻度处结合ADP分子,第二个刻度处俘获一个磷酸基团(Pi)并把它附着到ADP上,变成ATP,在生成这个化学键的同时锁定了能量(来自质子的滑落)。第三个刻度处打破结合键释放ATP分子。
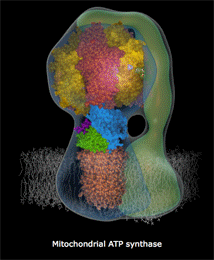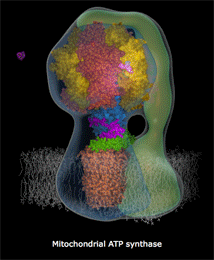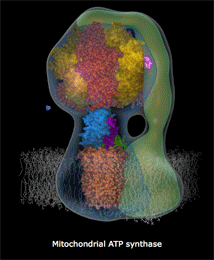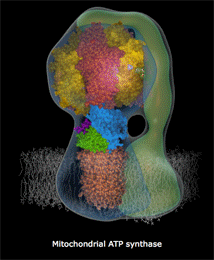
上面这个链接是精美的ATPase 精细结构动态图(下部模糊部分是线粒体内膜,H+从下方涌入),你可以看见整个机械的旋转,上部转子部分在每120度处蛋白结构形状和张力改变,ADP和Pi被“吸入”,ATP被释放。每个360度循环制造3个ATP。
人体内ATPase的旋转每秒大致150次,每天一个人的全身ATP更新量要超过自己的体重(当然这些分子都是循环使用的)。
这是一个自然界最微小的轮轴系统(“关于大自然为什么没发明轮子”的讨论可以休矣),一部细微精巧藐视所有人类技术的纳米机器。自己本身就是一条复杂而高效率的分子生产线。而且这个机器随时可以反向工作,即消耗内环境已有的ATP,把它们分解成ADP和Pi,利用释放的能量把质子泵向膜外(下一节会讨论什么情况下需要反向工作)。最初发现ATPase的时候,实际上发现的是它的后一种工作状态,也因此得名。让最初的发现者惊奇的是这个微小的酶工厂好像无所不在,所有真核生物的线粒体和叶绿体内膜,所有细菌的内膜上,都有基本相同的ATPase。当时的发现者说“ATPase的普遍,看起来就像生命的基本粒子”。远远在化学渗透发现之前,他根本没有意识到他说得有多么正确和深刻。
五 火星为什么是红色的,以及鞭毛公案
做基础科学研究的人,能遇到的最让人郁闷的问题就是这个:
“这个研究到底有什么实际应用?”
不去评价发问者暴露了什么样的思想境界,只说被提问者。很多基础科学研究者被这样问的时候,倾向于变得自我防卫。比如我前面说“相对论用在GPS校正上”,这就很防卫。其实那只是相对论最不值一提的后果。
我最喜欢美国一个理论物理学家的回答。当他代表美国的大型强子对撞机项目向政府申请十亿美元拨款时,国会要求他出席听证会,在听证会上他就被议员们这样问。他的回答是:“从现在已知的技术来看,绝对、绝对没有任何实际应用。”
“那么有什么理由花十亿美元去做呢?”
“因为我们想知道。其实你们也想知道。只是你们不知道你们想知道。”
申请失败了。但是这个回答却值得载入史册。绝大多数基础科学研究,都不是受实际的技术需要推动的。也不可能:既然是未知的东西,谁又能预测它的应用?整个科学都是靠人类旺盛的好奇心,靠人类天生对秩序、逻辑和知识的热爱推动的。科学是美丽神秘的森林,技术只不过是沿路采摘的果实。知识本身不但可以应用,而且会改变我们个人和社会看待整个世界的眼光和方法,从根本上使我们的存在变得更广大,更美好。
像物理、化学、生物这些基础学科,在一般人的眼中很可能枯燥且无实用价值,然而对于真正学懂而且融会贯通的人,他们眼中的世界跟我们普通人是不一样的。大概十年前,我有幸遇到这样一个人,在短短一小时内就给了我以上的感悟,让我发现了在一个锐利的头脑支配下,简单的科学原理可以变得如此明彻,充满力量。

那是在美国的一个小天文馆,当时有一群大学新生(大概是个兴趣小组)在搞参观活动,馆长出来接待(我是不沾边的,记住了答案的旁观游客)。馆长是一个穿着夏威夷衬衫的笑眯眯小老头,学生们连珠炮一样提问,他随口回答:
“火星为什么是红色的?”
“红色是三价铁的颜色—火星表面被氧化了。金星也是一样。铁锈是什么颜色?”
“火星是铁的吗?”
“任何岩石行星都可以说是铁的。铁是宇宙中最多的金属元素。”
“为什么?”
“铁的原子核结合能在所有元素中最低。恒星在燃烧,最终的灰烬都是铁。我们的行星都来自于灰烬。地球也不例外。地球有一个融化的大铁核。不然你认为地磁场是哪里来的?”
“那地球怎么不是红色的,怎么没有被氧化?”
“谁说不是?科罗拉多大峡谷看起来就像火星。其他的地方,我们有生命保护着。”
“火星上不是没有氧气吗?”
“没有,但是火星上曾经有水。”
“水到哪里去了?”
“太阳紫外线和宇宙射线把它烤干了。这是化学的烤干,在海洋的表面,微量的水被射线分解成氧气和氢气,火星大概用了20亿年。来,谁来给我写个反应式?”
“那火星上有很多氧气和氢气?”
“氧气和岩石中的铁结合了,变成了红色的氧化铁。还会氧化其他的岩石成分。氢气太轻,火星重力拉不住它,泄漏到宇宙中去了。”
“地球为什么没有被烤干?”
“因为地球有很多氧气。分解出来的氢气一离开水面,就和大气层的氧气重新结合,变回了水,最终会回到海洋。”
“为什么地球很很多氧气?”
“因为我们有光合作用,有生命。火星上氧气不能积累是因为微量的氧气被铁和碳不断消耗,固定在岩石中。地球上的固定氧被风和水不断粉碎,又被光合作用释放出来。”
“也就是说生命保护了地球?”
“是的,地球也保护了生命。”
“地球上为什么会有生命,而火星上没有?”
“这个,我不知道…”
小老头终于被问倒,全体学生大笑,我却一直在惊奇和震撼中。他说的全是简单物理、化学,我在学校基本都学过,却从来没有给我如此透彻的视界。
从那时起,我明白了科学=世界观。它不断回答我们对这个世界、对我们自身大大小小,乃至终极的疑问。下面要提到的公案,就是不同的回答在科学层面上的角力。
美国乃至整个西方,在二战后掀起了一股原教旨主义的潮流。如果你留意过近年美国社会的争论焦点和美国大选的论题,就会知道这不是一个小潮流,而是整个社会的精神大分裂。作为旁观的中国人来看,有的时候觉得美国人真是神经,总统辩论、政治风向天天吵这些鸡毛蒜皮的事。殊不知,这是两个阵营之间从世界观到价值观全方位的大战,这些鸡毛蒜皮都是刀尖。
新的原教旨主义者分成两种,一种在自己的地盘上以自己的武器战斗,比如坚称圣经句句真理。一种在现代文明的地盘上用科学的武器战斗,比如我们下面将要提到的鞭毛争论。比较好玩的是无论哪种论调,都很会给自己取名字。名字通常都冠冕堂皇充满现代知性的光辉,不管内容有多蒙昧。
比如“支持生命“(Pro Life)。什么意思?要求立法不准堕胎,极端者甚至不准强奸、乱伦的受害者堕胎。前几天共和党候选人罗姆尼宣布自己Pro Life,特别加上了“除了强奸、乱伦和危及母亲生命”,以示自己是温和派。与之相反支持堕胎权利的主张叫“支持选择(Pro Choice)”。
再比如“家庭价值”(Family Value)。什么意思?反对同性恋,以圣经中的判决来认定他们是社会的终极罪人。
还有“青年地球”(Young Earth)。这些人认为地球上有生命的历史不超过4000年。因为按照圣经上面的事件年代计算是这样的。比较牛X的青年地球专家,天天给公众解释诺亚方舟是怎么能够容纳一对霸王龙的。(霸王龙化石在美国太有名,你很难跟化石战斗)。

再比如“平等教育”(Equal Educational Opportunity)。什么意思?美国是宪法规定政教分离的国家,接受政府资助的学校都不能教任何宗教。“平等教育”论者要求你可以讲进化论,但必须用同样的课时讲“替代观点”。否则你就不准讲进化论。
什么替代观点呢?不,我们不讲经书,不呼上帝之名,讲“智慧设计”(Intelligent Design)。
智慧设计论者是原教旨主义者中的胆小鬼。他们千方百计回避直呼宗教之名,以绕过法律和舆论的障碍,而把矛头指向基督教最痛恨的科学:进化论。智慧设计论者也是外派的特种部队,他们通常都会用科学的语言(未见得是科学的方法)来发起挑战。比较老也比较经典的一个论战是关于眼睛的问题。智慧设计论者说,眼睛是一个非常精密的装置,各个部件离开整体都没有什么用处,因此它不可能是逐渐进化出来的(因为你们进化论者说进化出来的东西必然有用嘛!),那么,它只能是某种高级智慧设计出来的。什么高级智慧?可能我不知道。它的名字我不能说。这个逻辑,他们称作Irreducible Complexity, “不可降解的复杂度”。
关于眼睛的论战跟这篇文章的科学关系不大,只说结果:它被完全地驳倒了。生物学家找到了若干种中间形态和中间产物的生物证据。事实上这场论战推动了这方面研究的发展。
进入九十年代,智慧设计论者找到他们新的旗手:比希(Behe),以及新的战场:细菌鞭毛。比希是美国LeHigh大学的生物化学教授。我在第一节中曾把他归类于神棍,这里要致下歉,因为比希比通常的神棍要严谨、讲道理得太多太多。比希在2000年后,曾经出来纠正他以往的看法,说他不再相信“青年地球”,并相信生物有共同祖先。问题在于比希被自己的论点捆绑得太深(我最近在LeHigh大学的主页上还看见校方的免责声明:我们尊重比希教授的独立见解,但我们大学整体不同意他的任何理论。呵呵),而且他被原教旨主义的狂潮无限炒作、歪曲和滥用了。
比希的原始论点是这样的:很多细菌都有自己的运动装置,鞭毛。鞭毛是由几十个蛋白质部件组合而成的,各自有非常独立和确定的作用,使得细菌可以在液体中快速运动。这个机械的各部分分开都没有明确的作用,而且非常复杂,因此,这是不可降解的复杂度,因此,它不可能是进化来的,因此…简而言之,这就是眼睛问题的翻版,不同在于深入到分子生物化学的层面,显得异常高深和有说服力。1996年,他把鞭毛理论上升到对整个达尔文进化论解释的质疑,写成一本书:《达尔文的黑匣子》。这本书在美国超级畅销,远远超过一般高水平的科普书籍。一时间智慧设计论尘嚣直上,所有的宣传都鹦鹉学舌般应用鞭毛悖论。在google上搜“鞭毛”和任何智慧设计论关键词的组合,回来的结果都能淹死你。比希一炮打响,影响力到了这种程度:美国一场“平等教育”论者起诉学校的风头官司,从小地方直打到宪法法院,比希作为原告唯一的科学界证人出庭作证:智慧设计论到底是不是科学?结果官司是打输了。比希成了悲情英雄,这场官司也就成了美国政府为了党派利益压制“科学研究”的铁证,好比美国“平等教育派”的温泉关战役。
我们借助2000年左右的科学深度,来看看鞭毛到底是个什么东西。
鞭毛是一个复合蛋白装置,穿透细菌的细胞膜延伸到外部。在膜内部,鞭毛的蛋白构件呈同心圆环状排列,有三个环绕根基分开120度的蛋白“齿”,作为定子的凸起曲杆。鞭毛受化学渗透作用驱动,细胞膜外的质子水库透过穿透鞭毛的同心圆蛋白孔道滑落,赋予定子张力,定子顺次扭转一部分化学键,驱动中间的转子旋转。转子是穿透整个装置延伸到外部的蛋白构件,形状就像你喝可乐的麦秆,外部弯折,拐弯处还有一个蛋白护套。转子根部受定子驱动而旋转,外部跟着搅动,就像你用手搓麦秆,吸口处发生的搅动。细菌就用这样的装置“划水”,转子速度可达每秒几千转,细菌的运动速度可达每秒60个身位,(猎豹的最高速度也才每秒25个身位),可见这个运动机器的高效。鞭毛还有一个开关蛋白,稍稍变化形状就能反转整个装置旋转的方向。

确实是非常复杂精密的机器,难怪比希会产生那样的哲思。但是这个结构和驱动原理是不是似曾相识?
没错。比希在具体方法上的悲哀之处就在于他的书几乎和沃克的研究同时代,错过了沃克及其他人对ATPase的后续挖掘。还有一个悲哀是在他的时代,蛋白质基因定序分析来源的技术还没有成熟。不管是在结构上,驱动原理上,还是在组成蛋白的来源上,现在的分子生物学界都明确了ATPase是鞭毛的进化雏形。其中还有好几个中间状态。大致是这样一种进化阶段关系:
原始生命的胞膜进出孔道 --> ATPase --> III型细菌分泌系统 --> 细菌粘附装置 --> 细菌纤毛 --> 鞭毛
每个阶段的形态都受化学渗透驱动,采用一样的动力结构,而特化或者移植新的蛋白成分。其中“III型细菌分泌系统”是细菌特有的一种主动运输装置,它和鞭毛的相似是论战一开始就被其他科学家指出的。它含有一根由ATPase中空转子特化形成的“针管”,细菌用来分泌化学物质到环境中,给其他微生物“打针”,甚至用来和其他细菌“性交”,它也是鞭毛转子的雏形。
比希的“不可降解”理论在新的科学证据下跌得粉粉碎,曾经有一个反对者尖刻地说:“不可降解?是你不会降解。”但是直到今天,鞭毛仍然是智慧设计论者的“上帝之鞭”,你上网随便看一篇他们的宣传都能看见他们兴高采烈地挥舞。
化学渗透和ATPase在生物学理论上最根本的意义就是它完全是生命的基本特征。原核生物细菌、古菌类,真核生物的线粒体和叶绿体,全都使用相同的动力,全都拥有基本一致的ATPase。在最原始的细菌中,质子动力的普遍性和根本性表现得最充分,细菌不仅用化学渗透制造ATP,还用它进出口物质,驱动自己运动,保持膜外的质子浓度是细菌压倒一切的代谢优先任务。细菌是比我们高明得多的化学家,不仅仅使用氧气氧化糖来泵出质子(有时候还使用钠离子替代质子),还使用几乎是无限多种氧化还原反应,比如用硫化物氧化铁盐(这种代谢贡献了我们现在开采的所有的高品位铁矿带),或者硫化氢与甲烷,甚至还有氧化铀元素的强悍细菌(它们聚集的铀残留物曾经让法国人相信非洲国家偷偷研发了核武器)。能在生物体内成立的氧化还原动力,细菌几乎都发明了。不管是那一种化学动力,最终都归结于泵出质子。当氧化还原反应原料不够,膜外H+浓度告急时,很多细菌还停止其他一切活动,采用最古老的无氧呼吸:发酵,低效率地生产ATP,用这些ATP来驱动ATPase反向工作,把质子泵出去,保持膜外浓度—因为细菌需要化学渗透不仅仅是为了生产ATP,它是细菌的生命线,一切生命活动的基础。进化到你我这样的人类,这种机制仍然存在:你狂奔之后是否肌肉酸痛?那是因为你的体内的“细菌动力”—线粒体跟不上能量需求,细胞发酵葡萄糖产生半截子的乳酸和ATP,是乳酸的毒性让你刺痛。
我们可以放心地说,生命最普遍的特征就是生命最初的特征。化学渗透普遍、基本到这样的程度,它的发现对生命起源的理论必然造成重大影响。下一部分(也是最后一部分),将讨论化学渗透在生命起源上的意义。这里还要还比希先生一个公道:他的例证和论点虽然被粉碎了,但是他的内在逻辑并没有。因为我们已经看到化学渗透和ATPase本身也是非常复杂的机理和装置,那么在生命之初,他们又是怎么产生,怎么进化来的呢?这是科学现在还不能自信回答的问题,在这个问题上,各种理论的交锋,激烈程度不比美国政客玩堕胎差。不同的是,每种论点都有他自己的科学实证基础,交锋和证伪中大家的认识不断进化,而不是龟缩到2000年前地中海边的古书。
六 吹皱一池春水
生命起源的问题,向来是生物学中的皇冠,同时也是Mission Impossible。这是一个发生在近40亿年前,绝大部分物质证据都早已湮没的问题。又是一个我们已知的世界中只发生过一次的孤立事件,没有可以归纳的基础。同时它横跨生物、化学、地理、地质、气候、海洋、宇宙学诸领域,涉及的已知和未知千头万绪。最麻烦的是,即使有人真的发展出和事实相符合的理论,也几乎没有可能验证—这个实验本身很可能耗时千万年,或者上亿年。然而这丝毫没有驱散各领域科学家对这个问题无休止的兴趣。自从宗教的枷锁打破之后,科学性的生命起源假说有不下几十种,随着各学科的进步不断演进,不断丰富细节和淘汰伪说。
这些学说在方法上大体上分为两类,一类是正向的还原法,即不断搜集地球生命初始时代遗留下来的蛛丝马迹,包括化石、碳痕迹、同位素异常等等,不断试图还原当时的大气、海洋或者地质化学条件,试图通过模拟的实验,一点点验证生命起源的历程。这是传统的方法,它搜集和整理了大量的证据和数据,尤其在年代测定方面发展了很多高明的技术,取得了很多成果。到目前为止,最早的生命活动残迹可以上溯到38亿年之前,是由极其古老的岩石中碳同位素扰动间接证明的(生命是已知唯一改变碳同位素比例的自然过程)。
另一类是反向的反演法。最早的生命虽然早已死亡,但是它们的后代充满地球生物圈。这些后代种类极其繁杂,但是它们在分子生物和生物化学的层面具有令人惊奇的共性。这除了让科学研究者更加坚定了“生命同源”的信心之外,还提供了一个新的思路:共性即共祖特征。共性越多的生物,它们的亲缘关系越近。越普遍的共性,越有可能是生命在遗传树“根部”的特性。绝对普遍的生物共性,几乎一定是生物起源时的特性。跟传统的还原法正好相反(还原法是苦于证据太少太不完整),现代的生理学和基因科学为反演法提供了海量的各种生物遗传信息。整个生物圈的数码化遗传信息:基因组都是证据,只看研究者如何去挖掘和解读。难在如何穿透近40亿年生物分化、变异和灭绝造成的数据迷雾。如果我们能反推出最初的生物是什么样的特性,则如何起源的问题就解决了大半。因为在生物学中,“你是什么”基本就表述了“环境怎样造就了你”。反演法的研究最喜欢的一个术语就是LUCA(Last Universal Common Ancestor):“最后的共同祖先”。它是一个定义在逻辑上的生物,是现存所有生物的共同祖先。(之所以说最后,是因为不排除它之前更早的分支已灭绝且未留下后代)
曾经有一个建立在还原法上的假说,占据了权威地位,基本统治了二十世纪的大半部分。它的统治地位来源于50年代著名的“米勒实验”。米勒实验在简单的封闭系统中用氢气、甲烷和氨混合的气体模拟地球约40亿年前的“原始还原性大气”,辅以含水环境(模拟原始海洋)和不断制造的电火花(模拟大气中的雷电),在数天的时间内,就检测到了溶液中有基本有机小分子生成,时间越长数量越多,种类越丰富,包括各种氨基酸和核酸。这个实验当时可谓震惊科学界,因为谁也没有预计到如此简单的无机物原料和模拟环境,就制造出了蛋白质和DNA、RNA的基本组成单位,完成了从无生命到“几乎是生命”的飞跃。
有机小分子在原始海洋中不断生成,浓度越来越大,整个海洋成了富含有机物的“浓汤”。各种大分子(蛋白质、DNA、RNA)由小分子通过化学键像串珠一样串起来而生成(这是一个有实验基础的自发化学过程),开始互相竞争、淘汰、变异、组合。DNA、RNA的稳定性、自我复制性,辅以蛋白质的无限生物化学多变性使它们最终形成了机器-联盟,开始主动摄取能量,自我保存,自我传播—这已经是生命的定义。
这就是大名鼎鼎的“原始汤”假说。至少在我念书的年代,它还是各种学院教育中的正统学说,我相信现在不少国家的中学、大学课本仍然是这样教的。但以21世纪的研究眼光来看,这个假说已经被三振出局了。—不能怪其他领域的科学社区或者教育者们反应慢,因为并没有哪个学说在它之后取得那样的权威地位。

首先动摇它的根基的是地质学和太阳系研究的进展。米勒实验本身的初始条件,也是受更早的理论指导的,最早在20年代,英国的哈尔丹和俄国的欧柏林就同时提出了原始还原性大气的假说。他们当时的根据是:氢气、甲烷和氨,这是木星的大气成分(可以通过光谱技术精确测定)。地球和木星同样在太阳系的冷却中形成,那么假设地球初始大气和木星一样是有道理的。在米勒实验辉煌的光环下,这个假设维持了四十多年,直到60年代才被注意到它的粗陋。现在的看法是:一,地球初生年代长达5亿年的陨石大轰炸,早就把原始的“木星三气”轰光。二,地质证据表明当时的大气主要含氮气(来自轰击化学和火山活动)、二氧化碳(来自火山活动)、水蒸气和其他微量气体。这样,米勒实验的初始条件被挖空了一半。
1967年,“斯皮格曼的怪物”又给了原始汤理论重重一击。斯皮格曼是美国伊利诺伊大学的分子生物学家,他设计了一个实验,想用来确定自然选择条件下最小的遗传个体。他把一种非常简单的病毒从寄生细胞里分离出来,放在充满必要营养物(即各种有机小分子)和必要催化酶的溶液中自由生长。所谓病毒,就是蛋白质壳包裹的核酸序列,最接近“原始汤”主角的东西。这种病毒只有4500个碱基字节长,可谓极其简单。最初这些病毒正常复制,接着,有一些复制发生变异,丢失了基因。如果在寄生细胞中,病毒丢失基因意味着缺失某种功能,下场就是死亡。但在这个接近“原始汤”的充裕环境中,这反而成了优势:更短的序列复制更快,抢占资源更多,淘汰更长的对手。最终的结果是病毒逐渐退化成了只有220个字节的破碎残片,以疯狂的速度复制,充满整个培养液。这个实验不但其他科学家重复结果相同,更有普遍意义的是换成其他更复杂的病毒,结果仍然类似。
“斯皮尔曼的怪物”除了否定“原始汤”理论的发展性,还颠覆了很多生物学中的根本概念。从前认为“复杂化”和“进化”是生物的默认发展方向,现在省悟了根本没有任何预设方向,只随环境适应。对原核生物来说,简单化和退化甚至更有优势。
米切尔的化学渗透理论,以及化学渗透的绝对普遍性,基本宣告了原始汤理论的谢幕。同时把生命“能量”的一面重新放在聚光灯下。前面说过,由于20世纪基因科学的盛况,不管是公众还是生物学界,都对能量面有所忽视,把注意力集中在生命的信息面,所谓“太初有词”。“词”就是DNA承载的信息,信息支配结构和功能,支配新陈代谢,是遗传的本质和生命的意义。化学渗透理论说,不完全是这样。
最大的一个悖论就是:在开放的原始海洋中,任何聚集的化学物质浓度都趋于扩散、淡化。这是简单物理原则决定的。而我们现在知道,所有生物最基础的能量代谢形式都是基于浓度聚集的质子动力,都依靠生物膜。这就成了典型的“蛋鸡问题”。没有DNA上的信息,谁制造了生物膜?没有生物膜驱动的能量,就没有基本代谢,那么用什么来维持整个生命机器,制造蛋白质和复制DNA?如果有更基本的能量代谢形式来启动,那么我们为什么在现存的生物信息和生理中找不到它的遗迹?
这个悖论破坏了原始汤理论的设定场所和基本逻辑。也开启了从能量角度阐释生命起源之门,大量的新假说涌现出来。生命能量的一面现在必须和信息的一面紧紧纠缠在一起。比如说,病毒,从前定义成最简单的生命,也有很多看法怀疑它是最初的生命,而现在可以肯定病毒不是最初的生命,甚至被重新定义成“介于生命和非生命之间”的物质。因为病毒没有自己的能量代谢设施。它只含有信息,以及利用其他生物细胞能量代谢设施为自己服务的本事,而且通常是严格地针对某一类生物细胞。它是一种绝对寄生的东西,只能由有能量代谢能力的生物派生出来。它是活细胞的信息残片,是基因组的逃亡者,是不断移民、掠食甚至定居的基因流民。
而细胞的本源性被大大提高,不再被仅仅看做是基因们的豪宅。Compartmentalization: “封闭化”,成了生命最基础的要求。
介绍两个有趣的新假说。第一个不太为人所知,但很有个性:完全是从“封闭化”入手的。它假设生命起源的地点不是开敞的原始海洋,而是某种浅海湾边的的石灰岩浸润地带。这种地带的石灰岩由于水的侵蚀和地质结晶过程,内部形成了非常细微的疏松孔道结构,这就是天然的封闭空间。生物大分子可以由海水浸润流入,原始海洋中富含硫-铁氧化还原材料,而且这些结构的薄壁允许Na+ Ca+离子(质子的可替代品)的浓度聚集以及半通透性。换句话说,自然无机物的自组织性“免费”提供了“细胞膜”和一切能量代谢材料,最初的生物大分子只需要通过变异和选择发明利用的方法就够了。听起来够扯淡…但类似的地质结构确实在欧洲西北部的岛屿周围被发现了。而且实验表明,给定合适的浸润溶液配方,确实能够驱动化学渗透。只是如果想要测试生物大分子能在里面玩出什么花样,不知道要多少万年!
另一个是目前最火热的“黑烟囱”生命起源假说。它认为生命最可能的起源地点,是在深海底部高温、高硫、强酸性、无光、无氧的“黑烟囱”生态环境。这些地方是海底的地壳薄弱处,地热和高硫熔岩驱使高温海水从地底喷出,由于喷泉含大量的硫,而且出口逐渐沉淀成高达数十米的岩层,看起来就像冒着一股股黑烟的烟囱,因而得名。这种地貌现在在太平洋的海底就现实存在,而且拥有一个相当多样化的生物环境(甚至有些相当高等的动物也在这里定居,如一些蟹类,他们在这里适应得能忍受120度的高温,你休想煮熟它)。这也是现在世界上唯一一种不依靠光合作用提供基本能量的生态圈。这里的基础生物是古菌类,原核生物除了细菌之外的另一大基本门类,历史和细菌一样的古老。仅仅上面对黑烟囱环境的描述,就能看出化学渗透理论对这种假说的基本指导:白给的环境能量,白给的氧化-还原反应材料,严格地排除氧气(氧气对于原始生物都是剧毒的,学会用氧代谢是光合作用发明之后的事,但其他种类的氧化还原反应是基本需要),甚至白给的强酸性环境。如果生命起源于这种环境,只需要用着白给的能量发明膜就行了。

上图是一个典型的当代深海黑烟囱,这里没有光合作用,但有一个兴旺的生态系统。
另一类研究方法:LUCA模型的建立,更是压倒性地从能量生物学的角度入手。因为与能量代谢相关的基因,正是所有生物中最古老,变异最慢,共性最大也最核心的基因。生物学家们讨论和试图描述LUCA的时候,不是关注它长什么模样,有多大个头,而是关注它执行何种氧化还原反应,有哪些代谢酶,生活在什么样的地理和化学环境。这已经完全是一种崭新的体系了。
总之,九十年代之后的生命起源研究现状是群雄并起,天下大乱。理论更新速度之快,新发现新假说之层出不穷,都是二十世纪前面大半部分所无法企及的。固然是因为各个学科的知识积累都在推进,但化学渗透理论的建立,无疑是新纪元的开始。
讨论
评论
- 2015-12-08 10:18:14
刘小康这篇文章太棒了! - 2015-12-08 10:51:49
Jerkwin我们一起谢谢作者吧.
